2020年7月6日,美国约翰霍普金斯大学医学院肿瘤系Sidney Kimmel综合癌症研究中心的Jonathan D. Powell教授课题组在Nat Rev Cancer上发表了题为“Metabolism of immune cells in cancer”的综述 ,试图阐明癌细胞代谢和免疫代谢之间相互作用。作者认为通过了解包括对癌症的免疫反应在内的各种细胞的不同代谢需求,可以选择性调节免疫细胞功能,并对癌症和免疫新陈代谢的评估,进而揭示新陈代谢的治疗窗口并进行干预。

摘要
免疫检查点的封闭和过继细胞疗法的成功,使免疫疗法成为治疗癌症的成熟手段。细胞代谢已成为癌细胞和免疫细胞维持活力和功能的关键因素。为了维持肿瘤间庞大的合成代谢需求,肿瘤采用了与普通体细胞不同的代谢机制。这种代谢方式可以导致肿瘤微环境呈酸性,低氧,和/或耗尽了免疫细胞所需的关键营养物质。在这种情况下,肿瘤代谢本身就可以限制免疫检查点对肿瘤细胞的破坏。由于我们对免疫细胞代谢和癌症代谢的理解在过去十年中逐渐加深,我们希望能够阐明癌细胞代谢和免疫代谢之间相互作用。尽管有些代谢过程似乎是癌症和免疫细胞应答的基础,但代谢异质性和可塑性可能有助于区分两者。因此,了解包括对癌症的免疫反应在内的各种细胞的不同代谢需求,可以选择性调节免疫细胞功能。对癌症和免疫新陈代谢的评估可以揭示新陈代谢的治疗窗口并进行干预。
前言
过去几十年的研究表明,激活的免疫细胞可以利用多条代谢途径帮助癌细胞生长(图1)。这种代谢适应的融合使肿瘤微环境(TME)中的癌细胞和免疫细胞进行营养竞争。然而,我们发现癌细胞与免疫细胞的代谢程序之间以及不同免疫细胞之间存在根本差异。了解这些差异可以为发现针对新陈代谢开展肿瘤治疗提供支持。


癌细胞具有上调葡萄糖的糖酵解分解代谢,形成乳酸盐的能力,即使在氧气充足的条件下(有氧糖酵解),该效应被称为“ Warburg效应”,也被认为是恶性肿瘤的标志。现在人们越来越清楚地认识到,癌症的代谢也具有异质性,癌细胞可以参与各种代谢程序以满足生长和增殖的需求,除有氧糖酵解之外,线粒体呼吸也至关重要。我们可以发现具有高代谢活性的癌细胞(图1)会对TME产生深远的影响,导致营养物质消耗,缺氧,酸性和代谢产物的产生,这些代谢产物在浓度过高下可能具有毒性。来自TME的大量葡萄糖通过有氧糖酵解代谢,产生大量的乳酸和H +,从而降低了肿瘤内的pH。也就是说,产生乳酸的糖酵解和氧化磷酸化(OXPHOS)之间的平衡可能取决于缺氧的程度,缺氧的程度在TME中既可以是异质的,又可以是广泛的。值得注意的是,在中度低氧区域,线粒体呼吸产生的CO2被细胞外碳酸酐酶水合,形成HCO3-和H +。因此,氧化代谢可能是TME中细胞外酸化的重要且易被忽视的来源。
最近的免疫疗法包括免疫检查点的抑制与过继细胞疗法(CAR T)在肿瘤免疫中得到广泛应用。例如,黑色素瘤细胞中糖酵解代谢的增加与对过继性T细胞疗法和免疫检查点的抗性相关。其他研究表明,肿瘤细胞上的免疫检查点(包括PD1和B7-H3)发出的信号导致TME中的葡萄糖消耗增加。有趣的是,实际上肿瘤酸化的直接结果就是诱导了免疫抑制检查点相关途径。此外,免疫检查点的阻断可以抑制肿瘤细胞的糖酵解,恢复TME中的葡萄糖,并允许T细胞糖酵解和细胞因子的产生。最近的一些研究表明,针对肿瘤固有代谢的特定方面,例如六胺生物合成途径(HBP)或谷氨酰胺代谢,可以促进免疫反应并使肿瘤获得对免疫检查点封闭的敏感性。
由于免疫治疗已成为肿瘤治疗的支柱,因此深入了解浸润性免疫细胞与癌症之间的代谢依赖性变得越来越重要。本综述旨在讨论以下基本问题:哪些代谢程序对于参与癌症免疫应答的细胞亚群至关重要;这些代谢程序在TME中如何受到干扰;TME中代谢紊乱对当前免疫治疗的影响;以及如何利用代谢干预措施来增强抗肿瘤免疫反应。
肿瘤免疫微环境
癌细胞所特有的高代谢途径(图1)对TME中营养成分等影响较大,并可能对免疫反应产生关键影响。癌细胞的高代谢活性和TME中紊乱的脉管系统都可以导致营养物质的缺乏和低氧微环境,在癌细胞与浸润的免疫细胞之间建立代谢竞争。在小鼠模型中,抗肿瘤CD4 + T细胞的葡萄糖摄取和效应功能与癌细胞的糖酵解活性成反比。此外,对来自TCGA的黑色素瘤患者的转录组分析显示,效应T(Teff)细胞基因,例如CD40lg和IFNG,与HK2(糖酵解通路的限速酶)表达呈反相关。在TME细胞内活跃的代谢程序也可能导致某些代谢产物的毒性浓度增加。TME中已报道了腺苷,犬尿氨酸,鸟氨酸,活性氧(ROS)和钾的水平升高,以及酸中毒增加都会在抑制抗肿瘤免疫反应方面产生深远影响。
TME的免疫环境由一系列不同的细胞类型构成(表1)。效应细胞执行细胞杀伤功能,无论在天然免疫还是适应性免疫反应中。在适应性免疫反应中的抗肿瘤细胞包括CD4 +和CD8 + Teff细胞,它们通过不同机制杀死癌细胞。CD8 + Teff细胞通过诱导凋亡和细胞因子分泌直接杀死肿瘤细胞中。CD4 + T细胞包含许多细胞亚群。其中一些细胞亚群(如Th1)也具有显着的抗肿瘤活性。这些抗肿瘤CD4 + T细胞(CD4 + conv)与免疫抑制性CD4 + T细胞(Treg)不同。尽管CD4 + conv细胞可能参与直接杀死肿瘤细胞,但它们主要通过细胞因子分泌和协助CD8 + T细胞活化来促进抗肿瘤免疫。抗肿瘤CD4 + conv T细胞与CD8 + Teff细胞具有相同重要的代谢特征。尽管就抗肿瘤免疫性而言,B细胞的研究较少,但B细胞也可能在TME中发挥效应作用。重要的是,作为适应性免疫系统的一部分,T细胞和B细胞会产生记忆细胞群,这些记忆细胞群在感染或肿瘤反应消退后会持续很长时间。CD8 +记忆T细胞(T mem)是长期肿瘤控制的关键。先天细胞,例如自然杀伤(NK)细胞和炎性巨噬细胞,也具有关键的抗肿瘤效应功能。TME内还有免疫抑制细胞群,包括CD4 + FOXP3 + Treg细胞,髓样抑制细胞(MDSC),抗炎巨噬细胞和一些B细胞亚群。抗原呈递细胞,例如肿瘤内树突状细胞(DC),在维持TME内的适应性免疫反应起重要作用。

抗肿瘤反应的代谢
抗肿瘤效应T细胞的葡萄糖代谢
CD4 + conv和CD8 + Teff细胞形成了抗肿瘤免疫反应的关键细胞群。幼稚的T细胞在识别抗原提呈和共刺激因子的信号后,可以进行增殖以及代谢。尽管许多早期研究说明有氧糖酵解的上调是T细胞活化的标志,但现研究表明三羧酸(TCA)循环代谢和OXPHOS上调也是CD4 + conv和CD8 + T细胞活化的关键。尽管TCA循环代谢在激活后24 h内被上调,但有氧糖酵解上调更为迅速,发生在激活后6 h内。
MYC和缺氧诱导因子1(HIF-1)的转录活性均受T细胞活化而上调,并促进代谢重编程。值得注意的是,尽管HIF-1调节缺氧反应的代谢,但在缺氧条件下,其活性也会因响应T细胞活化而被诱导。MYC和HIF-1的转录活性导致编码促进糖酵解的酶(例如丙酮酸激酶(PKM1),己糖激酶2(HK2)和GLUT1)相关基因的表达上调。糖酵解途径中来自近端代谢产物的途径也是T细胞活化和功能的组成部分(图1)。磷酸戊糖途径(PPP)代谢6-磷酸葡萄糖生成NADPH和5-核糖。在CD4 + T细胞激活后,进入PPP的葡萄糖显着增加。PPP是NADPH的主要细胞来源,也是新激活的CD8 + T细胞中脂肪酸和质膜合成所必需。NADPH对于REDOX稳态在增殖哺乳动物细胞中也至关重要。激活的T细胞中的ROS水平需要进行精细调节。尽管失调的ROS水平可能具有毒性,但ROS在Teff细胞活化中起着重要作用,已被证明可促进CD4 +和CD8 + T细胞中NFAT依赖性的IL-2表达。另一条起源于早期糖酵解反应的途径是HBP,是糖基化底物的主要细胞来源,它可介导多种蛋白质,包括稳定性,运输以及功能作用。HBP依赖于葡萄糖和谷氨酰胺的代谢,并且对它们发生反应。由HBP产生的主要底物UDP-GlcNAc对于效应CD4 +和CD8 + T细胞的扩增和功能至关重要。最后,丝氨酸-甘氨酸-一碳途径使细胞能够生成用于叶酸循环的丝氨酸,甘氨酸,NADPH和一碳单位。Teff细胞的增殖和功能在体外和体内都取决于丝氨酸的充分代谢(图1)。
未代谢为乳酸或通过糖酵解途径代谢的葡萄糖碳对Teff细胞中的TCA循环有重要作用6(图1)。在高度增殖的细胞中,TCA循环的中间体被迅速消耗,以作为广泛的生物分子合成的基础,这一过程称为 cataplerosis。例如,柠檬酸盐可以输出到细胞质中以再生成乙酰辅酶A,用于脂质和胆固醇的合成,这两者对于在增生的Teff细胞中产生细胞膜至关重要。像癌细胞一样,Teff细胞具有高度增殖能力,并上调特定的糖酵解程序,包括有氧糖酵解,PPP,HBP和TCA循环,以实现大量的细胞分裂和效应功能。
TME中的T细胞和葡萄糖受限
TME中的葡萄糖受限会明显影响T细胞反应。例如,低葡萄糖条件(0.1μmM)抑制了T细胞中糖酵解中间体磷酸烯醇式丙酮酸(PEP)的生成,从而破坏了体外钙依赖性NFAT信号传导。与对照相比,降低生长培养基中的葡萄糖浓度可以抑制细胞外酸化率,增加耗氧率,减弱mTOR信号传导并抑制两者的效应功能。降低的mTOR复合物1(mTORC1)信号干扰Teff细胞分化,并且在CD4 + T细胞的情况下有利于免疫抑制性Treg细胞的发展。有趣的是,在CD8 + T细胞中,雷帕霉素对mTOR的阻滞促进了Tmem细胞的分化,这可能在维持抗肿瘤反应中发挥重要作用。与对照培养基相比,葡萄糖的下降抑制了Teff细胞中关键效应分子干扰素-γ(INFγ),IL-17和颗粒酶B的产生。原发性卵巢癌细胞条件下培养基中的葡萄糖限制导致microRNA介导的组蛋白甲基化酶EZH2的抑制,导致NOTCH信号减少,细胞因子产生受到抑制以及Teff细胞活力降低。
在共培养实验增加小鼠肉瘤细胞的糖酵解能力,可以导致CD8 + T细胞效应功能受到抑制。同样,与野生型肿瘤相比,在小鼠模型中,植入过表达Hk2的黑色素瘤细胞抑制了CD4 + T细胞的抗肿瘤效应功能和体内反应。此外,黑色素瘤和非小细胞肺癌患者肿瘤样品中糖酵解相关基因的表达与T细胞浸润呈负相关。我们也可以通过直接操纵T细胞代谢来实现新陈代谢平衡。例如,与对照相比,在黑色素瘤中肿瘤特异性CD4 + T细胞中糖酵解酶PEP羧激酶的过表达可以改善抗肿瘤反应。
线粒体呼吸也是Teff细胞代谢的关键。最近的研究报道,癌症患者的T细胞(与健康对照相比)和荷瘤小鼠的肿瘤浸润性CD8 + T细胞(与非浸润性CD8 + T细胞相比)显示线粒体质量降低以及线粒体功能障碍。与健康对照相比,慢性淋巴细胞性白血病患者外周CD8 + T细胞的线粒体受损。此外,这些患者对CAR T细胞疗法的反应程度与注入的CAR T细胞的线粒体损伤程度呈负相关。与健康供体的CD8 + T细胞相比,肾细胞癌患者的肿瘤浸润性CD8 + T细胞显示线粒体动力学和功能失调,包括线粒体ROS水平升高和超极化。这些T细胞的正常体外活化可以通过线粒体ROS清除剂或丙酮酸补充进行挽救。线粒体的生物发生和功能在功能失调的肿瘤浸润性CD8 + T细胞的亚群(特别是耗竭性T细胞)中极为紊乱。总体而言,这些研究表明癌症本身可以导致Teff细胞代谢紊乱,包括线粒体动力学,并且癌细胞的糖酵解活性程度与浸润性T细胞的抗肿瘤效应功能之间存在相互关系。
氨基酸与抗肿瘤T细胞反应
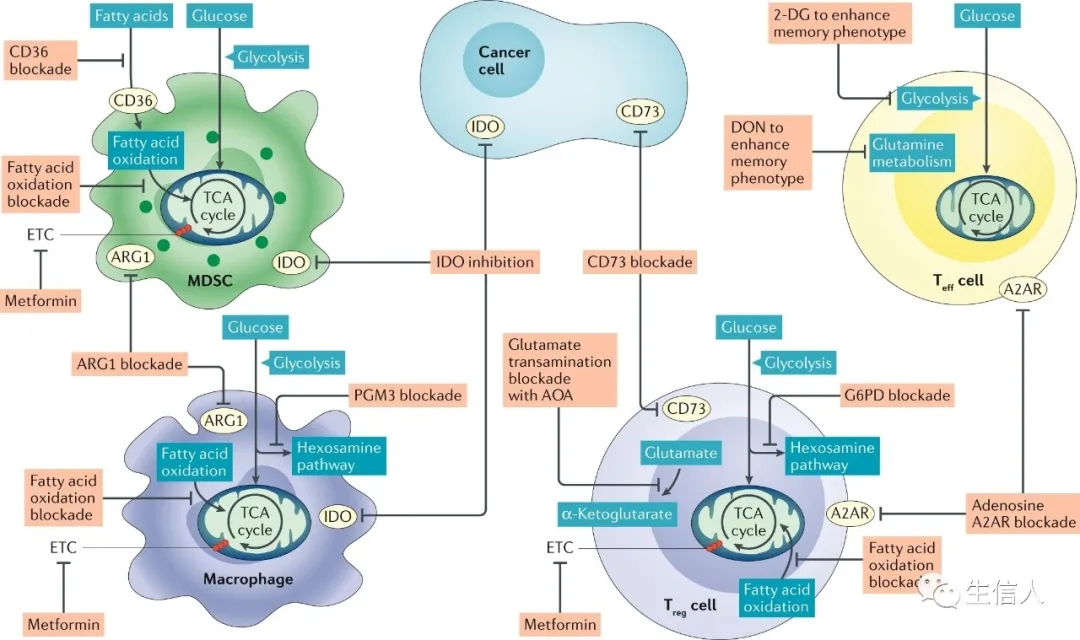
像癌细胞一样,高度增殖的免疫细胞(例如活化的T细胞)也依赖于氨基酸代谢来支持蛋白质和核苷酸的合成。因此,氨基酸转运蛋白,包括SLC7A5(也称为LAT1),SLC38A1(也称为SNAT1),SLC38A2(也称为SNAT2)和SLC1A5(也称为ASCT2)在T细胞激活后被高度上调。必需氨基酸必须是外源获得。例如,亮氨酸是效应CD8 +和CD4 + conv T细胞中mTORC1信号传导,效应功能和分化所必需。活化的T细胞还可以快速代谢精氨酸,外源性精氨酸的补充可以改善T细胞的适应性并增加中央Tmem细胞的生成。丝氨酸,色氨酸和半胱氨酸也是T细胞反应的重要营养成分,因此是抗肿瘤免疫反应的重要介质。色氨酸是必需氨基酸,在TME中是决定T细胞反应强度和有效性的重要因素。与正常培养基相比,在不含色氨酸的培养基中,人类T细胞的增殖和活化受到了强烈抑制。癌细胞,肿瘤相关巨噬细胞(TAM),MDSC,抑制性DC和癌症相关成纤维细胞可通过吲哚胺2,3-二加氧酶(IDO)的酶促活性耗尽色氨酸水平,这些酶可在TME内高水平表达。代谢途径对肿瘤生长的十分重要,IDO表达与多种癌症类型(包括胃癌,结肠直肠癌,非小细胞肺癌和黑色素瘤)患者的不良预后相关。
在增殖细胞中,谷氨酰胺为氨基酸和核酸的合成提供氮,为TCA循环中间体提供补充的碳,这个过程被称为“ anaplerosis”(图1)。癌细胞和一些活化的免疫细胞,例如T细胞和巨噬细胞,通常富含高度谷氨酰胺。在体外刺激鼠CD4 + conv T细胞的过程中,谷氨酰胺转运蛋白SLC1A5和SLC38A1和/或SLC38A2的表达显着上调。在MYC的驱动下,谷氨酰胺被谷氨酰胺酶(GLS)代谢为谷氨酸,在由谷氨酸脱氢酶(GLUD1)转化为α-酮戊二酸(αKG;也称为2-氧戊二酸)后,谷氨酰胺可能进入TCA循环。随后在TCA周期中,αKG被代谢为琥珀酸盐和富马酸盐。有趣的是,在谷氨酰胺受限的情况下,一些癌细胞系转换为葡萄糖引起的 anaplerosis,其中丙酮酸被丙酮酸羧化酶转化为草酰乙酸,进入TCA循环。最近表明,在体外谷氨酰胺阻断的条件下,效应CD8 + T细胞也能够上调丙酮酸羧化酶的活性。
尽管通过限制培养基中的谷氨酰胺可以抑制分化的CD8 + Teff细胞的效应功能和增殖,但如果在激活CD8 + T细胞的过程中限制了谷氨酰胺的利用率,它会改变向CD8+记忆表型的分化,研究表明分化作用是由αKG介导的。αKG和其他TCA代谢物(例如琥珀酸酯和富马酸酯)可以调节多种细胞过程的活性,包括表观遗传重塑和关键转录因子(如HIF-1α)的稳定性。
脂质代谢与T细胞
活化的T细胞还可以重新编程脂质代谢,上调脂质合成和胆固醇摄取,其对细胞膜合成至关重要,并分别由转录因子固醇调节元件结合蛋白1(SREBP1)和SREBP2介导。在缺少SREBP1和SREBP2功能的活化小鼠CD8 + T细胞中,增殖,代谢重编程和抗病毒活性被显着抑制。此外,在体外CD8 + T细胞活化和扩增过程中,细胞膜中的胆固醇含量部分受胆固醇酯化酶乙酰辅酶A乙酰转移酶(ACAT1)调节。在过继转移小鼠肿瘤模型中,Acat1基因敲除的CD8 + T细胞表现出增加的膜胆固醇和改善的T细胞受体聚集和信号传导,从而导致增殖,功能增强和杀伤力增强。
用阿瓦西米对ACAT1的药理抑制作用改善了小鼠的抗肿瘤作用。最新研究 证明肿瘤中高胆固醇可通过激活内质网应激反应而诱发T细胞功能障碍。因此,尽管胆固醇对于Teff细胞的增殖和代谢很重要,但针对胆固醇代谢特定方面改善抗肿瘤免疫反应的益处尚需进一步研究。
免疫记忆的代谢
与Teff细胞不同,CD8 + Tmem细胞优先依赖OXPHOS。与CD8 + Teff细胞相比,增强的备用呼吸能力也是Tmem细胞的高度特征。最初研究显示使用埃托莫西酯作为肉碱棕榈酰转移酶1A(CPT1A)的抑制剂,该酶是线粒体转运蛋白,负责长链脂肪酸的进入以进行脂肪酸β-氧化(FAO),表明FAO是Tmem中OXPHOS的主要燃料。然而,最近使用T细胞特异性Cpt1a敲除模型进行的研究对此提出了质疑,并证明了高剂量依托莫昔(200μm)的脱靶效应可能是原因。
TCA循环的中间体,例如αKG,琥珀酸酯和富马酸酯,在适应性记忆中特别重要。通过改变这些TCA代谢产物抑制2-氧戊二酸依赖性双加氧酶(2OGDD)已显示可增加CD8 + T细胞中的记忆细胞分化。尽管葡萄糖,谷氨酰胺和脂肪酸是推动TCA循环的主要营养来源,但一系列其他营养物质(例如氨基酸和乙酸盐)也可以进入该循环。特别是,乙酸酯代谢正在成为CD8 + T细胞和某些癌症类型中乙酰辅酶A的重要来源。在线粒体中,乙酸盐在被酰基辅酶A合成酶短链家族成员1(ACSS1)代谢形成乙酰辅酶A后可以进入TCA循环。另外,乙酸盐可以在细胞质中通过ACSS2转化为乙酰辅酶A,在乙酸中它可以促进脂肪酸合成和乙酰化反应,这对于表观遗传重编程和翻译后修饰有重要意义。乙酸盐的代谢是促进记忆CD8 + T细胞功能的重要代谢途径。有趣的是,在T细胞活化过程中,谷氨酰胺代谢的阻滞增加了Tmem细胞的分化,并诱导了乙酸代谢和相关的酶,包括ACSS1和ACSS216,85。作为静止细胞,Tmem细胞相对于有氧糖酵解优先依赖于OXPHOS,并且具有显着的线粒体储备,这是抗原活化后进一步上调OXPHOS所必需。Tmem细胞可以采用几种不同的营养来源,以促进这种代谢程序。
缺氧与抗肿瘤T细胞反应
尽管肿瘤具有高度异质性,但高水平的代谢活性和相关的氧气消耗以及功能紊乱,功能差的脉管系统仍会产生缺氧区域,中位氧饱和度水平低于2%(正常组织中值约为5%) 。缺氧对Teff细胞的影响并不直接。在常氧条件下,HIF-1转录活性会响应T细胞活化而被上调,因此要了解缺氧对进一步增强HIF-1活性的影响,同时还要评估HIF-1的依赖效应,具有一定的挑战性。早期对CD8 + Teff细胞活化,分化和功能的体外研究表明,尽管在缺氧条件下抑制了增殖和某些细胞因子的表达,但其溶解能力,活化标记物和存活率却得到了改善。随后的体内研究表明,与暴露于环境O2张力(20%)的小鼠相比,暴露于低于大气O2张力(8%)的小鼠中CD4 +和CD8 +脾T细胞的活化更弱。其他研究表明,体外低氧暴露会导致代谢物(S)-2-羟基戊二酸(S-2-HG)在细胞内积聚,从而深刻改变CD8 + T细胞的活化和分化,抑制细胞因子的分泌和溶细胞能力。但有趣的是,在小鼠模型体内转移后增强增殖,长期生存和抗肿瘤反应。糖酵解活性通过组成性HIF-1α活性(通过有条件地敲除HIF-1调节剂Vhl来实现)实际上促进了疫苗模型中长效效应记忆细胞的数量。其他工作表明,缺氧诱导了TME中各种细胞外切核苷酸酶CD39和CD73的表达,这些酶将TME中的ATP分解为腺苷。腺苷是A2A和A2B嘌呤能受体的配体,它们在广泛的免疫细胞中表达,并且广泛地具有免疫抑制作用,抑制效应细胞功能和Teff细胞增殖。有趣的是,补充氧气通过下调腺苷信号传导途径来增强小鼠T细胞的抗肿瘤免疫反应。缺氧对抗肿瘤T细胞的影响是一个不断发展的研究领域。考虑到肿瘤中低氧区域的普及以及低氧对适应性免疫反应的深远影响,其进一步的研究将显然有益于免疫治疗领域。
毒性代谢物
除腺苷外,癌细胞代谢产生的许多其他产物也会影响浸润的T细胞(图1和2)。在小鼠和人CD8 + T细胞的体外研究中,TME中细胞外乳酸和H +的水平升高可以抑制T细胞增殖,存活,细胞毒性和细胞因子的产生。与标准生长培养基相比,在高水平的乳酸和H +存在下,小鼠CD8 + T细胞的体外激活过程中,编码关键Teff细胞转录因子NFAT的基因的上调受到损害。体内小鼠研究表明,与空载细胞相比,被Ldha敲除的小鼠黑色素瘤细胞产生的乳酸更少,对免疫介导的肿瘤排斥反应的反应更强。与对照培养基相比,在乳酸和H +升高的情况下激活的人效应CD8 + T细胞中,MAP激酶信号传导也受到严重破坏。

肿瘤中特定氨基酸的积累也可以抑制Teff细胞反应。在这方面,研究得最好的是色氨酸代谢产物,尤其是犬尿氨酸,其作用是通过IDO1的活性产生。与对照相比,犬尿氨酸在体外引起活化CD8 + T细胞上PD1协同抑制途径的上调。与对照相比,在用外源性犬尿氨酸处理的小鼠模型中,在肿瘤浸润的CD8 + T细胞上也观察到PD1的这种上调。来自经犬尿氨酸处理的荷瘤小鼠的肿瘤浸润CD8 + T细胞产生的IFNγ和TNF较少。
在报道说明癌细胞通过释放竞争性代谢物(R)-2-羟基戊二酸酯(R-2-HG)抑制T细胞活性。这种代谢物可以抑制表观遗传的双加氧酶,例如组蛋白脱甲基酶,从而导致甲基化增强和转录修饰。由异柠檬酸脱氢酶(IDH)突变的人脑胶质瘤产生的R-2-HG在体外研究中被T细胞吸收。R-2-HG在体外干扰活化CD4 +和CD8 + T细胞的增殖,T细胞受体信号传导,NFAT活性和多胺生物合成。在IDH突变的神经胶质瘤小鼠中R-2-HG释放入TME抑制补体介导的抗肿瘤反应以及T细胞迁移,增殖和细胞因子分泌。这些研究突出了TME内癌症代谢物和免疫功能之间的复杂相互作用(图2)。
高度坏死导致TME中钾的含量增加,从而限制了T细胞效应功能。该状态由降低的乙酰辅酶A胞质水平介导,此状态诱导了活化T细胞的表观遗传重塑,从而导致TME中Teff细胞功能异常。但是,这种功能障碍状态富含T细胞干性的特征。高钾浓度的Teff细胞在离体刺激和扩增后产生的T细胞具有更高的体内持久性,多能性和肿瘤清除能力。通过肿瘤坏死和TME固有的代谢活性产生T细胞抑制性代谢产物,是逃避肿瘤免疫的重要机制。
代谢和天然效应反应
由于NK细胞在主要组织相容性复合物I类(MHC-I)下调(癌细胞的常见逃避策略)期间杀死细胞,所以它们形成了先天反应的关键效应组成成分。在体外,NK细胞的细胞因子刺激(IL-12和IL-15)后,有氧糖酵解和OXPHOS在代谢上被上调。有趣的是,在体外NK细胞刺激过程中,这些细胞因子诱导的代谢变化需要SREBP转录因子。在体外,SREBP活性的药理抑制作用抑制了代谢重编程,细胞因子产生和细胞毒性,并在过继性NK细胞小鼠模型中减少了抗肿瘤反应。有趣的是,据报道内源性SREBP抑制剂(例如27-羟基胆固醇)可在TME中增加,因此可能是NK细胞抑制的机制。小鼠的肺癌进展和与肿瘤相关的转化生长因子-β(TGFβ)与肿瘤相关的NK细胞中果糖-1,6-双磷酸酶(FBP1)表达的增加有关。FBP1是糖异生的关键酶,一旦激活,FBP1会强烈抑制NK细胞的糖酵解,从而导致功能障碍和活力降低。有趣的是,FBP1的药理抑制作用足以在体外重建糖酵解代谢以及细胞因子的产生和细胞毒性,并改善过继细胞疗法小鼠模型的抗肿瘤反应。这些研究表明,通过FBP1抑制来挽救NK功能取决于葡萄糖代谢的恢复,因为用2-脱氧葡萄糖(2-DG)阻断葡萄糖代谢阻止了FBP1抑制引起的拯救。2-DG本身也导致NK细胞功能异常,这表明抑制葡萄糖代谢可能对NK细胞抗肿瘤反应产生深远影响。TME内的其他代谢紊乱也可能影响NK细胞功能。例如,精氨酸水平低会损害NK细胞的增殖和IFNγ的产生,而缺氧会抑制细胞溶解活性。在体外研究中,人类NK细胞激活受体(例如NKp46和NKp30)在响应低氧或精氨酸水平较低时受到抑制。如在TME中一样,高乳酸水平和相关的低pH值在体外研究中也抑制了NK细胞的细胞毒性,细胞因子产生和NFAT信号传导。最后,TME中腺苷水平升高可强烈抑制NK细胞效应功能和增殖。
其他先天细胞,巨噬细胞和DC也会在激活时引发特定的代谢程序。在早期的体外研究中,将巨噬细胞分为炎性(M1)或免疫抑制(M2)表型,但很少有证据表明这些极化表型在体内发挥着独特的作用。最近发现了以不同转录状态为特征的一系列巨噬细胞表型。也就是说,具有炎性特征的巨噬细胞在抗肿瘤免疫中发挥重要作用。葡萄糖代谢是巨噬细胞中炎症表型的重要方面。被Toll样受体激动剂激活后,这些细胞显示出糖酵解基因的表达增加,葡萄糖摄取的高水平,乳酸的产生增加以及谷氨酰胺过表达。这种代谢重编程导致琥珀酸水平升高,从而通过稳定HIF-1而增加了炎性细胞因子IL-1β的表达。炎性巨噬细胞还特别依赖于PPP来产生NADPH,13C-葡萄糖示踪研究证实,通过激活培养物中的炎性表型,通过该途径的葡萄糖路径增加。NADPH对于产生高水平的ROS(作为氧化性爆发的一部分)是必需的,这是这些细胞的关键效应机制。精氨酸也是促炎性“ M1”巨噬细胞功能中的关键营养物质,因为在体外研究中,精氨酸比其他活化或“ M2”极化巨噬细胞表达更高水平的诱导型一氧化氮合酶(iNOS)。iNOS需要精氨酸才能产生细胞毒性一氧化氮,这是抗肿瘤反应的重要促炎介质。
TME中特定的营养缺乏,特别是葡萄糖和精氨酸,会严重限制这些细胞中新陈代谢和效应程序。葡萄糖限制不仅可以整体抑制糖酵解,而且可以降低PPP活性和TCA循环功能,从而限制了NADPH,ROS和琥珀酸酯的生成,所有这些都可以严重限制M1巨噬细胞的功能。2-DG的糖酵解抑制作用显着减少了巨噬细胞促炎细胞因子的分泌。
DC是参与抗肿瘤应答的一类重要的抗原呈递细胞,能够进行抗原交叉呈递的肿瘤内DC已作为这种反应的重要组成部分。激活后,DC会成熟并允许抗原加工并呈递给T细胞。该反应与从OXPHOS到有氧糖酵解的代谢转换相关,其由HIF-1α响应体外LPS激活,并由PI3K-AKT途径响应Toll样受体体外刺激介导。在DC激活过程中,这种转换为糖酵解作用并远离OXPHOS的过程对于DC存活,刺激性细胞因子的产生以及T细胞的激活至关重要。有趣的是,AMPK的药理学激活可促进线粒体的生物发生和氧化呼吸,并足以在体外阻止DC成熟。鉴于这种对有氧糖酵解的关键依赖性,TME中的葡萄糖竞争可能会显着抑制DC激活和生存能力,从而限制DC促进有效和持久性T细胞反应的能力。
癌症免疫逃逸的新陈代谢
适应性免疫抑制的代谢
免疫抑制性Treg细胞优先依赖TCA循环和线粒体呼吸。尽管初步研究表明Treg细胞对FAO的依赖性并未说明依托莫昔的脱靶作用,但其他研究表明,FAO确实支持Treg细胞中的OXPHOS,尽管不是唯一途径。与Teff细胞相反,Treg细胞在体外显示降低葡萄糖摄取并表达较低水平的GLUT1的表达。有趣的是,尽管糖酵解似乎并未在Treg细胞分化或长寿表型中发挥关键作用,但据我们发现,一部分高活性Treg细胞(称为效应Treg细胞)依赖于糖酵解的上调以获得最佳功能。Treg细胞似乎具有新陈代谢的灵活性,可以在相对苛刻和异质的条件下(例如TME)生长。为此,据报道,Treg细胞转录因子FOXP3通过抑制MYC(有利于OXPHOS和NAD(H)氧化)来重新编程细胞代谢。在低葡萄糖和高乳酸的条件下(例如在TME中发现),这些适应作用使这些免疫抑制细胞具有代谢优势,从而使Treg细胞能够在体外抵抗乳酸诱导的功能性和增殖性抑制(不同于Teff细胞)。在体外实验中,培养基中的葡萄糖或谷氨酰胺缺乏(导致细胞内αKG减少)可以改变CD4分化并促进Treg细胞的发育。
与Teff细胞相似,Treg细胞对缺氧的反应尚不完全清楚。缺氧已显示可促进细胞因子介导的Treg细胞募集进入肿瘤环境。FOXP3转录本实际上是响应HIF-1α诱导而增加。此外,与野生型对照相比,过继转移的Treg细胞特异性Hif1敲除细胞在小鼠模型中未能迁移到脑肿瘤中,在用二氯乙酸酯处理的Treg细胞中也观察到糖酵解受到抑制。有趣的是,在混合淋巴细胞培养实验中,通过Treg细胞上腺苷受体A2A的低氧反应性腺苷信号传导诱导了增殖,并显着增强了免疫调节活性。有文献报道低氧诱导的HIF-1α可以破坏Treg细胞的稳定性,并表明低氧可以通过直接HIF-1α与分别定义细胞亚型的转录因子FOXP3和RORγt的直接相互作用来促进TH17 CD4 + T细胞。
TME中氨基酸的独特代谢也可以对Treg细胞产生深远的影响。IDO1活性可以在体外强烈促进Treg细胞分化,这种作用似乎是色氨酸缺乏症以及下游代谢产物(如犬尿氨酸)的产生所致。
天然免疫抑制的代谢
TAM表型为高度免疫抑制。像Treg细胞一样,M2巨噬细胞上调FAO和线粒体呼吸。尽管早期研究表明FAO必须在M2巨噬细胞中发挥作用,但并未考虑到CPT1A抑制剂etomoxir的脱靶作用,但由于过表达Pgc1α引发巨噬细胞而导致免疫抑制表型,并且抑制促炎细胞因子的产生。M2巨噬细胞以不同于炎性巨噬细胞的方式代谢氨基酸,表达高水平的精氨酸酶1(ARG1),这会消耗精氨酸并生成多胺,多胺是伤口愈合的重要介质,但也具有高度免疫抑制作用。
另一组与肿瘤相关的免疫抑制先天细胞MDSC似乎具有高代谢活性。与周围的MDSCs相比,与肿瘤相关的MDSCs中有氧糖酵解和OXPHOS均上调。在另一项研究中,与来自相同小鼠的脾嗜中性白细胞相比,来自荷瘤小鼠脾脏的粒细胞MDSCs也显示出有氧糖酵解和OXPHOS的增加。有趣的是,在小鼠乳腺癌模型中,MDSC的体外扩增和在TME中的蓄积可通过用2-DG阻断糖酵解来减弱,这很可能是由于这些细胞中的ROS水平升高所致。
肿瘤中的缺氧区域与巨噬细胞的积累有关,它们在巨噬细胞中通过产生与肿瘤转移有关的血管生成因子,促有丝分裂因子和细胞因子来协助肿瘤的发展。此外,缺氧可促进免疫抑制巨噬细胞表型的产生。缺氧可产生腺苷,可通过巨噬细胞上的A2A和A2B受体触发信号传导,这两者均可增强免疫抑制巨噬细胞的分化和功能能力,并减弱体外炎性巨噬细胞的细胞因子释放。已证明培养物中乳酸的升高可促进M2表型,增加ARG1表达和多胺依赖性免疫抑制。在三阴性乳腺癌中,较高的糖酵解率显示出促进MDSCs,而在这些癌细胞中限制糖酵解则抑制了癌细胞中粒细胞集落刺激因子和粒细胞-巨噬细胞集落刺激因子的分泌,并限制了MDSC的发育。有趣的是,与常氧性MDSCs相比,低氧使MDSCs趋向于具有免疫抑制性的M2样TAM表型。这是通过HIF-1α机制引发,因为在小鼠黑素瘤肿瘤模型中,与野生型MDSC相比,Hif1a基因敲除的MDSCs显示出肿瘤生长增加。
利用不同代谢的可塑性
虽然可以通过抑制许多代谢途径来减弱Teff细胞的激活,增殖和功能,但是其他属性,例如重新刺激时的长期生存力或效应功能可能增强。尽管用2-DG抑制糖酵解代谢抑制了Teff细胞的产生,但它也使T细胞趋向于长效记忆表型。有趣的是,在再次输注用于肿瘤治疗之前,在离体T细胞活化和扩增过程中阻断糖酵解,不仅可以提高抗肿瘤T细胞的存活率,还可以改善细胞因子的产生和细胞毒性。此外,有研究证明了该途径在维持谷氨酰胺阻断的CD8 + T细胞中维持代谢稳态方面的重要性。这些发现可能表明一种普遍的治疗策略,即阻止使用典型的代谢燃料(例如葡萄糖或谷氨酰胺)可能使某些癌症在代谢上受到损害,但鉴于抗肿瘤性T细胞使用其他来源的能力,它们可能会在代谢方面保持完整和正常运转,如醋酸盐。尽管可以通过药理学方法将特定的代谢干预作为检查点封闭的辅助手段引入,但这些靶点可能特别适用于CAR T细胞疗法,其中可以通过遗传手段精确定义代谢途径。
免疫检查点封闭与免疫代谢
定义检查点疗法的代谢后果和其反应后的代谢情况都非常有意义。在多项研究中,检查点信号已显示出调节代谢的作用。例如,癌细胞上PDL1的表达可以驱动癌细胞中Akt-mTOR的活化和糖酵解,增加葡萄糖的摄取并增强与T细胞竞争葡萄糖的能力。人胃癌组织中T细胞中的CD155-TIGIT信号传导抑制了葡萄糖的摄取,乳酸的产生以及糖酵解酶GLUT1和HK2180的表达。相反,与同种型处理的对照T细胞相比,共刺激途径GITR的激动作用广泛提高了T细胞的代谢活性和增殖。最后,活化的人T细胞上的体外PD1和CTLA4信号抑制了与T细胞活化相关的代谢途径,如有氧糖酵解。为此,将代谢抑制剂与检查点抑制剂结合使用的前景有望提高检查点封闭的疗效。同样,通过对T细胞进行代谢重编程以使其更强大,持久的记忆细胞可能会改善其对检查点抑制剂的反应。最近的临床试验预示了抗叶酸培美曲塞与抗PDL1免疫检查点封闭的结合。除具有直接的抗肿瘤作用外,培美曲塞治疗还增强了抗肿瘤CD8 + T细胞的代谢适应性和效应功能,并诱导了癌细胞的免疫细胞死亡,从而触发了免疫反应。
结论与展望
尽管免疫代谢的许多基础已从癌症代谢的观察中获悉,但很明显,癌症与免疫代谢重编程之间存在明显差异。这些差异为靶向代谢提供机会,以此作为增强免疫疗法功效的手段(图3)。这种方法可以通过许多不同的策略来实现。这些措施包括靶向肿瘤代谢程序以抑制生长和改变TME,靶向抑制性免疫细胞的代谢以抑制其功能以及靶向效应细胞代谢以增强肿瘤杀伤力。同样,在过继细胞治疗之前,T细胞代谢途径的体外药理或基因重编程也可以增强其功能,包括延长寿命或增强效应功能。
未来工作应集中于TME内免疫细胞和癌细胞的代谢相互依赖性。除了营养消耗和可以抑制一定浓度下的免疫反应的代谢产物产生之外,癌细胞还可以与TME中的其他细胞发生代谢扰动,其中可以诱导并共同的代谢程序,从而有利于恶性进展。据报道,胰腺星状细胞可以为癌细胞提供丙氨酸,并且已经报道了骨髓基质细胞提供半胱氨酸以促进慢性淋巴细胞性白血病细胞的存活。在另一份报告中,来自癌细胞谷氨酰胺代谢的氨通过TME扩散并触发了与癌症相关的成纤维细胞的自噬,这反过来又提供了蛋白质分解产物,例如谷氨酰胺本身,以进一步支持癌细胞代谢。重要的是要了解逃避免疫的癌症是否以及通过何种机制选择免疫细胞的代谢机制,并从其显着的代谢灵活性中受益。

 京公网安备
11010802036025号
京公网安备
11010802036025号